Man-Seok Bang1,
Chang Rae Rho2,
Bong-Hui Kang3,
Kyong Jin Cho1,4 ![]() ,
Chung-Hun Oh1,5
,
Chung-Hun Oh1,5
For correspondence:- Kyong Cho Email: choh@dankook.ac.kr Tel:+82415501918
Received: 18 October 2016 Accepted: 10 February 2017 Published: 01 April 2017
Citation:
Bang M, Rho CR, Kang B, Cho KJ, Oh C.
Overex
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the ex
Methods: Twenty pterygium and 20 healthy conjunctival samples were obtained from 19 patients who consented to pterygium excision and conjunctival autograft surgery. Tissues (pterygium and normal conjunctiva) were analyzed using next-generation RNA sequencing. Significant gene lists were obtained using DAVID, GSEA, and KEGG for enriched pathway analyses of differentially expressed genes. Real-time polymerase chain reaction was performed to validate differential ex
Results: There was 3.15 ± 2.6, 3.21 ± 4.6, 3.03 ± 1.83, 4.28 ± 6.12, and 4.96 ± 1.89-fold increase in the ex
Conclusion: Alternative and lectin complement systems are activated in pterygia samples compared to normal conjunctival samples
Introduction
Pterygium is an overgrowth of fibrovascular tissue, with a winglike appearance, from the conjunctiva over the cornea [1,2]. Although several theories on its pathogenesis have been postulated, including immunological mechanisms, infection, and ultraviolet exposure [3-5], the definitive cause of pterygium is still not known. Some studies reported that immunological mechanisms such as type 1, 3, and 4 hypersensitivities might contribute to its pathogenesis, and large numbers of infiltrating lymphocytes, predominantly T-cells (CD3+), have been found in the substantia propria of pterygium specimens [6,7].
The complement system is a part of the immune system that boosts the ability of antibodies and phagocytic cells to clear bacteria and damaged cells from the host. Complement activation products mediate immune reactions and the expression of many inflammatory cytokines [8-10], and the complement pathway has been related with the pathogenesis of many diseases. Variants in several genes, such as complement 2 (C2), complement 3 (C3), complement factor H (CFH), and complement factor B (CFB), encoding complement pathway proteins, have been associated with age-related macular degeneration [11].
The classical pathway is triggered by activation of the C1-complex. The C1-complex is composed of C1q, C1r and C1s, or C1qr2s2. This occurs when C1q binds to IgM or IgG complexed with antigens. The binding of C1q to a ligand results in a conformational change which leads to the sequential activation of C1r and C1s [12]. The alternative pathway dose not require a specific protein for the initiation stage. Spontaneous hydrolysis of C3 leads to constitutively low-grade complement activation [13]. The lectin pathway is initiated by the recognition of carbohydrate molecules (MBL, N-acetyl-glucosamine, mannose, fucose or glucose, and ficolins) on the surface of viruses, bacteria, and other pathogenic organisms [14]. MBL-associated serine proteases 1 and 2 are complexed with MBL and are stimulated upon MBL binding to sugar moieties, subsequently cleaving C4 and C2 to form the classical pathway of C3 convertase, C4b, and C2a [15].
Next-generation sequencing is a technique that parallelizes the sequencing process, allowing concurrent high-throughput genome sequencing. To characterize the immunopathology of pterygium, we used this next-generation RNA sequencing methodology to identify genes associated with pterygia that were not expressed in healthy conjunctival samples.
Methods
Patients and samples
The present study was reviewed and approved by the Institutional Review Board of Dankook University Hospital, Yongin, The Republic of Korea (code: 2016-04-016), and it adhered to the tenets of the Declaration of Helsinki [16]. Twenty pterygium and 20 normal conjunctival samples were obtained from 19 patients who underwent pterygium excision and conjunctival autograft surgery after obtaining their written informed consent.
RNA isolation and sequencing
Total RNA was obtained from pterygium and normal conjunctival samples using an RNeasy Micro kit (Qiagen, San Diego, CA, USA) according to the manufacturer's instructions. Total RNA quality and quantity were confirmed using a NanoDrop 2000 spectrometer (Thermo Scientific, Waltham, MA, USA) and Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA). To build Illumina-compatible libraries, we used a TruSeq RNA library preparation kit (Illumina, San Diego, CA, USA) according to the manufacturer's instructions. TruSeq RNA libraries constructed by polymerase chain reaction (PCR) amplification were quantified using quantitative PCR (qPCR) according to the qPCR Quantification Protocol Guide, and their quality was assessed using a Bioanalyzer 2100 (Agilent Technologies). The amplified library was sequenced using a HiSeq™ 2000 system (Illumina).
Raw read filtering and alignment with a human reference genome
Raw read filtering using Trim Galore (Babraham Bioinformatics, http://www.bioinformatics.babra ham. ac.uk/projects/trim_galore) used the default value to remove the reads with sequence adaptors and the low-quality reads. All subsequent analyses were based on the clean reads. The reference sequences were downloaded from the University of California at Santa Cruz genome browser (version hg19, https://genome.ucsc.edu/cgi-bin/hgGateway?db= hg 19). Trimming reads were respectively aligned to the reference genome using RNA-STAR (https://github.com/alexdobin/ STAR).
Analyses of differential expression and functional annotations
Reads uniquely mapped to a gene were calculated using the Cufflink package according to the default values. The gene expression level was expressed as the number of uniquely mapped fragments per kilobase of exon region per million mappable reads. Genes differentially expressed between paired pterygium and normal conjunctival samples were determined based on values of p ≤ 0.05 and fold change ≥ 2. Significant gene lists were generated using DAVID (Database for Annotation, Visualization, and Integrated Discovery, https://david.ncifcrf. gov/), GSEA (Gene Set Enrichment Analysis http://software.broadinstitute.org/gsea/index.jsp), and KEGG (Kyoto Encyclopedia of Genes and Genomes, http://www.genome.jp/kegg/) to produce functional annotation of differentially expressed genes. DAVID is a bioinformatics tool that systematically extracts biological meaning from large gene lists. GSEA generated a ranking based on the correlation of genes differentially expressed between pterygium and normal conjunctival samples. KEGG is a computational description of biological systems, consisting of the molecular structure of genes and proteins, integrated with information on molecular pattern charts of interaction, reaction, and association networks.
cDNA synthesis and real-time analysis
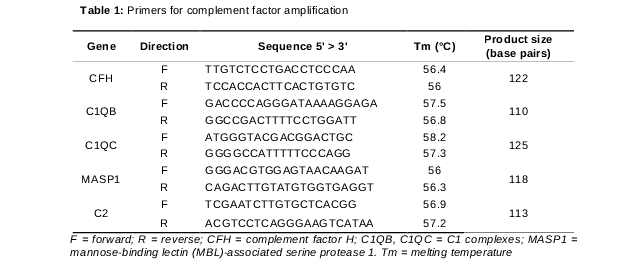
The cDNA synthesis was performed using an iScriptTM cDNA synthesis kit (Bio-Rad, Hercules, CA, USA) according to the manufacturer’s instructions. Real-time PCR (RT-PCR) was performed using the StepOne Plus real-time PCR system (Applied Biosystems, Foster City, CA, USA) with SYBR® Green PCR Master Mix (Applied Biosystems) according to the standard manufacturer’s instructions. Primers were purchased from Bioneer (Daejeon, The Republic of Korea). Primer sequences are listed in .
Evaluation of the pterygium area of the cornea
Photographic images of Anterior segment were imported into Adobe Photoshop CS5 (Adobe Systems, San Jose, CA, USA). Borders of the pterygium and cornea were marked, and the “pixel counter” tool was used to quantify and compare pixel counts in the digital image. The size of pterygium was calculated based on the pixel ratio of pterygium to a circular reference marker (cornea).
Statistical analysis
Statistical analysis was performed using SPSS statistical software for Windows, version 23.0 (SPSS, Chicago, IL, USA). A paired t-test was used to compare the expression of complement factors between pterygium and normal conjunctival samples. Pearson’s correlation analysis was used to determine possible correlations between pterygium size and the expression of complement factors. A value of p < 0.05 (two-tailed) was considered statistically significant.
Specimens
The mean age ± SD of 19 patients (10 females, 9 males) was 59.9 ± 7.4 years. Seventeen of the 20 collected pterygium samples were primary pterygium samples, and the remaining three samples were recurrent pterygium samples.
Results
RNA sequencing results
We studied the differentially expressed genes using RNA sequencing in eight pterygium samples and their matched healthy conjunctival samples. As a result, we generated an average of 55,550,533 reads (range: 43,439,664–80,389,513 reads). Also, we used the Spliced Transcripts Alignment to a Reference (STAR, http://pipe-star.readthedocs.io/en/latest/explain_ star.html) to map the reads to a human genome (UCSC hg19, http://hgdownload.cse.ucsc.edu/ downloads.html), thus obtaining an average value of 93 %.
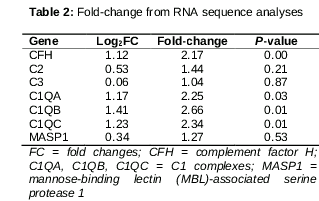
By comparing the transcriptome sequences of the pterygium samples and their matched conjunctival samples, several differentially expressed transcripts were identified. Furthermore, 917 genes were differentially expressed with a fold change ≥ 2 and p < 0.05, and among them, there were 641 upregulated genes and 276 downregulated genes. We performed future analyses of the differentially expressed gene functions using DAVID, the Gene Set Enrichment Assay, and the KEGG pathway.
As a result, we found that CFH, C1QA, C1QB, and C1QC were differentially expressed between pterygium and matched conjunctival samples () and they were clustered in the complement system.
Validation of complement factor overexpression using RT-PCR
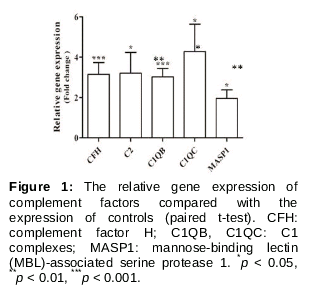
Quantitative real time PCR (qRT-PCR) was performed for confirming the relationship of the complement system genes with pterygium. As a result, complement factor genes (CFH, C2, C1QB, C1QC, and MASP1) were upregulated more than 2-fold. Notably, CFH and C1QB genes increased more than three-fold change and showed significant expression changes (p <0.001) compared to those in normal conjunctival tissue ().
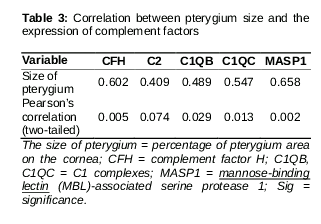
Relationship between gene expression and pterygium size
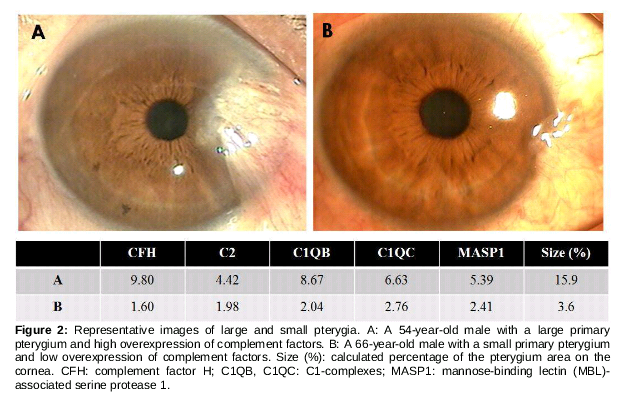
The range of the pterygium in the cornea was the measurement obtained for confirming the correlation between complement-related genes and pterygium size in 19 patients. As a result, the size of the pterygium increased when complement-related genes overexpressed compared to those in matched normal conjunctival tissues (, A). Conversely, when complement-related genes were relatively less expressed, the size of the pterygium was small (, B).
Discussion
Next-generation RNA sequencing has increased ourability to identify specific proteins involved in many diseases [17,18]. In the present study, this technique was used to identify putative differences in pterygia samples when compared with normal conjunctival samples. We observed that several genes were found to be overexpressed, or underexpressed in pterygia tissues when compared with matched normal conjunctival tissues. To the best of our knowledge, this is the first study to compare samples of pterygia and healthy conjunctivas using next-generation RNA sequencing. The results provide many new opportunities to study the causes and diagnoses of pterygium.
RT-PCR assays were performed to validate the overexpression of complement factors CFH, C2, C1QB, C1QC, and MASP1. We tried to determine the expression pattern of all complement pathways, including the classic pathway (C2, C1QB, and C1QC), the lectin pathway (C2 and MASP1), and the alternative pathway (CFH). The results showed that all of these complement factors were significantly increased in pterygia samples compared to normal conjunctival samples.
The complement system, a protein cascade involved in immune response, consists of several soluble and cell membrane-associated proteins [19]. This cascade is a part of the immune system that enhances the ability of antibodies and phagocytes to clear bacteria and damaged cells from an organism. The biochemical pathways that activate the complement system consist of the classical complement pathway, the alternative complement pathway, and the lectin pathway [20]. The classical pathway is stimulated upon binding of C1q to antibody-opsonized target cells. The lectin pathway is activated by mannose-binding lectin (MBL). The alternative pathway is induced by infectious agents including bacteria, viruses, and fungi. All pathways converge at the level of C3 to initiate the terminal pathway, which leads to the formation of C5b-9. In addition, anaphylatoxins, C3a and C5a, recruit immune cells to the site of inflammation [21].
To avoid damage to surrounding tissues, both soluble and membrane-bound regulators tightly regulate the complement system. The C1 inhibitor binds to C1r, C1s, and MASP-2 to prevent further cleavage of C4. C4b-binding protein inactivates the classical C3 convertase pathway and factors H, and I inhibit the alternative C3 convertase pathway by cleaving C3b in the enzyme complex [21]. CFH is a key regulator of the complement system in innate immunity that maintains the optimum C3 level in circulation [22]. Genetic alterations, including absolute factor H deficiency, have been associated with membranoproliferative glomerulonephritis and hemolytic uremic syndrome [23]. In our study, CFH was overexpressed in pterygia tissue compared to normal conjunctival tissue, but C3 expression was not increased. This might be the result of regulation by CFH.
We found that complement factors are overexpressed in pterygia samples compared to normal conjunctival samples. However, the magnitude of the changes differed among patients. We tried to identify the factors associated with these variations. Using Pearson’s correlation analyses, the size of the pterygium was correlated with the amount of complement factor expression. The occurrence and growth of pterygium are associated with immunological reaction and inflammation [6]. Therefore, large size of the pterygium might induce more severe activation of the complement pathway. This implies that the amount of complement factor reflected the severity of the pterygium and that, in clinical practice, detection of gene expression changes could be used to predict post-surgical lesions in the pterygium. However, the exact role of complement factors in the pathogenesis of pterygium is still not completely understood. The pathogenesis of pterygium resulting from ultraviolet (UV) light damage, oxidative stress, and microbial infection might activate the complement system such as the classic pathway (C2, C1QB, and C1QC) and the lectin pathway (C2 and MASP1). UV radiation in sunlight are the prime cause of skin cancer in humans, with both UVA and UVB wavelengths being important in photo-carcinogenesis. UV radiation has various biological effects on carcinogenesis [24]. Some of these effects, as well as other forms of stress, may activate the complement system [24] and cause chronic inflammation of the conjunctiva and limbus, resulting in initiation and progression of pterygium. Recent evidence suggests that potentially oncogenic viruses such as human papillomavirus and Epstein-Barr virus may be involved in the pathogenesis of pterygia [25]. Infection might be related to activation of the alternative pathway (CFH) and the lectin pathways (C2 and MASP1) in the pathogenesis of pterygium. The alternative and lectin pathway are induced by exposure to infectious agents including bacteria, viruses, and fungi.
Conclusion
The alternative and lectin complement systems are activated in pterygia samples compared to normal conjunctival samples. CFH is also overexpressed in pterygium samples. Pterygium size correlates with the amount of complement factor expression. Complement factors could be used as diagnostic and prognostic markers of pterygium.
Declarations
Acknowledgement
References
Archives


News Updates